パパイン様プロテアーゼ(パパインようプロテアーゼ、英: papain-like protease、PLpro)は、コロナウイルスの複製に不可欠なシステインプロテアーゼで、CAグループペプチダーゼ(C16ファミリー)に属する。パパイン様プロテアーゼ(PLpro)の主な機能は、ウイルスのポリタンパク質を協調して処理することである。ただし、PLproには、コロナウイルスの回避を助けるために、宿主細胞のタンパク質からユビキチンとISG15を除去するという付加的な機能がある。これらの理由から、PLproはSARS-CoVに対する抗ウイルス剤の研究における治療標的となっている。
分子構造と特徴
分子構造
SARS-CoVのパパイン様プロテアーゼ(PLpro)は、316個のアミノ酸モノマーで、4つの異なる構造ドメインから構成されており、「広げた右手」構造を形成している。そのうち3つのドメインはサム(親指)、パーム(手のひら)、フィンガー(指)ドメインと呼ばれ、4つ目のドメインはユビキチン様ドメイン(UBL)と呼ばれている。PLproのカルボキシル末端にはSUD(SARSユニークドメイン、SARS Unique Domain)が続き、アミノ末端にはNAB(核酸結合ドメイン、Nucleic Acid Binding)が続く。これらはPLproに隣接するドメインで、SARS-CoVの非構造タンパク質のゲノムやプロテオームに属しているが、PLproには含まれていない。具体的には、PLpro複合体は、213 kDaの膜関連タンパク質であるSARS-CoV nsp3(非構造ドメイン3、non-structural protein 3)のアミノ酸1541と1856の間に位置している。
PLproポリペプチド鎖の最初の62アミノ酸がUBLドメインを形成する。これは他の3つのドメインとは別のN末端ドメインで、ユビキチンタンパク質と非常によく似た二次構造をとっている。サムドメインは4本のαヘリックス(α4-7)で、パームドメインは6本鎖βシート(β8-13)で、フィンガードメインは4本鎖のねじれた逆平行βシート(β4-7)で構成されている。SARS-CoVのPLproの全体的な構造は、SARS-CoV-2のPLproと83%一致することが示されているが、いくつかの違いがある。例えば、UBLドメインのアミノ酸数は62個ではなく60個であり、サムドメインは4個ではなく6個のαヘリックス(α2-7)で構成されている。しかし、一方で、酵素の活性部位のすべての重要な残基は、PLproの両方の形態間で不変である。
PLproの活性部位は、システイン、ヒスチジン、アスパラギン酸からなる共通の触媒三残基で構成されている。システイン残基(C112)は、サムドメインのα4ヘリックスの基部にあり、パームドメインの基部にあるヒスチジン残基(H273)から3.7 Å離れたところにあり、BL2(G267-G272ループ)として知られる柔軟なループに隣接している。このループは6アミノ酸から構成されている。また、アスパラギン酸残基(D287)は、ヒスチジン残基から2.7 Å離れている。
フィンガードメインには、4つのシステイン残基を持つ2つのβヘアピンも見られる。これらの残基は、四面体の幾何学結合を亜鉛イオンと調整し、亜鉛結合サブドメインを形成する役割を果たす。亜鉛結合を形成するPLproのこの能力は、プロテアーゼ活性とその構造の完全性の維持に不可欠である。
一般に、PLproの三次構造は、USP7やUSP14などのUSP(Ubiquitin Specific Proteases、ユビキチン特異的プロテアーゼ)と非常によく似ている。
特徴
PLproは、ポリペプチド鎖中に多数のシステイン残基(約3.5%)を持つ、わずかに塩基性の易水溶性で高反応性のプロテアーゼである。すなわち、主鎖を構成する316残基のうち、11残基がシステインである。これらのうち、1つは活性中心の触媒システインであり、他の4つは亜鉛イオンとの結合を調整する役割を担っている。さらに、触媒システイン(C112)は、触媒三残基の中で最も反応性の高い残基であると考えられ、プロテアーゼの構造的および機能的コンホメーションにおけるこのアミノ酸の重要性は明らかである。
酵素の活性部位は、他の種類のPLproや、触媒システインを持つ他のプロテアーゼとも非常によく似た化学的性質を持っている。ほとんどの場合、触媒システインは求核性残基として働き、ヒスチジンは酸塩基対に類似し、アスパラギン酸はシステインの脱プロトン化を促進する。
作用機序
コロナウイルス(CoV)の複製は、ウイルスゲノムの完全な保護を常に意味する、複雑な機構が関与する高度に洗練されたプロセスである。複製タンパク質の合成は、ORF1a(オープンリーディングフレーム1a)とORFabがリボソーム機構によって翻訳されることで開始され、pp1a(486 kDa)とpp1abという2つのポリタンパク質が生成される。この2つのポリタンパク質は、ウイルスにコードされた2つのシステインプロテアーゼであるパパイン様プロテアーゼ(PLpro)と3-キモトリプシン様プロテアーゼ(3CLpro)によって処理される。PLproドメインは、非構造タンパク質3(nsp3)の一部としてコードされ、非構造タンパク質1および2を処理する。一方、3CLproはnsp5の一部であり、残りの12種類の非構造タンパク質を処理する。これらのnspの生成が完了すると、nspは宿主の膜上でウイルス複製複合体を組み立て、ウイルスゲノムの複製と転写を開始する。
触媒メカニズム
SARS-CoV PLproは、システインプロテアーゼの触媒サイクルを通じて機能する。このサイクルでは、Cys112が求核剤として機能し、His273が酸塩基機能を担い、最後にAsp287がヒスチジンとペアになってシステインの脱プロトン化を形成して促進される。
Cys112のプロトン化が基質への求核攻撃に先行するかどうかは、科学界でまだ分かっていない。また、反応性のある求核種がチオール-ヒスチジンのチオールイオンなのか、または、リガンドと反応後にチオニウムイオンが生成されるセリンペプチダーゼと同様のメカニズムで作用する中性電荷のチオールなのかについても不明である。
最初のステップ「E」では、SARS-CoV PLproの触媒残基は水素結合によって結合しており、比較的短い距離で分離されている(図を参照)。この事実は、プロトン化したCys112がHis273と平衡状態にあり、一方で、基質との結合によって反応性チオールが平衡状態になるという可能性を示す。しかし、基質が結合して複合体「ES」を形成する際に、Cys112の脱プロトン化が起こる可能性は否定されていない。次のステップでは、Cys112チオールがペプチド結合のカルボニル-カーボネートと反応し、負に帯電した四面体の中間体が形成される。この四面体の中間体は、トリプトファン残基(Trp107)を持つオキソアニオンで構成されており、生化学的基質が結合する酵素PLproの活性部位にある隣接するオキソアニオンホールの存在によって安定化されている。アスパラギン(Asn110)などの他の残基は、コロナウイルスのPLP2に共通して存在し、オキソアニオンホールの安定化にも寄与している。次のチオエステル中間体「F」は、C末端のアミドの除去と、ペプチド結合の破壊により形成される。チオエステルのカルボニル炭素に水分子が加わると、負に帯電した第2の四面体中間体「TI-2」または「FQ」が形成される。サイクルのこの時点で、PLproの活性部位にあるオキソアニオンホールによって、「TI-2」オキソアニオンの安定化が再び起こる。第5段階では、四面体の中間体からシステインが切断され、N末端の「EQ」カルボン酸が形成される。このカルボン酸は、形成された酸のカルボニル炭素とTrp107の窒素との間の水素結合を介して、PLproの活性部位に一時的に配置される。最後に、切断されたペプチドのN末端を除去することで、PLproの触媒サイクルが完了する。最終生成物「Q」が活性部位から放出され、その結果、酵素「E」が再生される。
PLproとUbおよびISG15の関係
PLproの触媒ドメインの分子構造は、「広げた右手」がN末端で「ユビキチン様」ドメイン(UBL)に結合して構成されている。PLproの構造は、脱ユビキチン化酵素(DUB)の構造と似ている。さらに、PLproの酵素活性のin vitro特性評価によって、このタンパク質様プロテアーゼはユビキチン(Ub)およびUBLのISG15(インターフェロン誘導遺伝子15、interferon-induced gene 15)タンパク質を認識して、加水分解することがわかった。
UbとISG15はどちらも、ウイルス感染に対する宿主の自然免疫系の免疫応答のシグナル伝達において重要な役割を果たしており、これらの欠如はウイルス増殖に有利に働くことが分かっている。
部位
SARS-CoV nsp3は、RNA複製に関与する213 kDaのポリペプチドであり、nsp3aからnsp3gまでの7つのドメインから構成されている。
nsp3のN末端領域は、ほとんどのコロナウイルスで完全に保存されている。nsp3dは、pp1aとpp1abを処理するパパイン様プロテアーゼ(PLpro)である。この領域には、球状のユビキチン様フォールド(ubiquitin-like、Ubl)に続いて、グルタミン酸に富むな酸性ドメインと、nsp3bにはADP-リボース-1-ホスホターゼドメインが含まれている。
SARS-CoV PLproの触媒ドメインとnsp3は小胞体の膜に位置しており、ほとんどのドメインは細胞質に存在する。このドメインは、複数の触媒活性のある酵素、膜貫通型ドメイン、および機能がまだ解明されていないドメインに隣接して見られる。
医学的重要性
急性呼吸器症候群(SARS-CoV)を引き起こす最初のコロナウイルスは、2002年に中国の広東省で発生した。この事実は、同ウイルスの感染症に対する治療法を研究するきっかけとなった。しかし、SARS-CoVパンデミックでは、8500人以上が感染し、推定約800人が死亡し、世界経済に数十億ドルの損失を与えたにもかかわらず、SARS-CoV感染症を治療するための臨床的に証明された抗ウイルス剤、ワクチン、モノクローナル抗体療法はまだ存在していない。
一方、コロナウイルスの複製に不可欠なパパイン様プロテアーゼ(PLpro)タンパク質は、その二重の機能性から、ウイルスの複製を阻害し、感染症の致死率を低下させる抗ウイルス剤の開発における重要な標的である。この観点から、ほとんどの研究は、SARS-CoV PLproの構造と機能、およびコロナウイルスの複製複合体の生合成におけるPLproの重要な役割の詳細な理解に向けられている。
さらに、SARS-CoV-2とSARS-CoV PLproには多くの類似した特徴があることから、SARS-CoV PLpro阻害剤の研究を進めることは、COVID-19に対して可能な治療法を研究する上での確かな出発点になると考えられる。
SARS-CoV PLpro阻害剤
一般に酵素阻害剤とは、酵素に結合してその活性を低下させる分子のことであり、SARS-CoV PLproの場合、このタンパク質の阻害剤として機能する可能性のあるさまざまな産物や化合物を見つけるために、いくつかの研究が行われてきた。各阻害剤は、IC50値(値が低いほど、酵素に対する阻害剤の効力が高い)によって酵素との親和性を特徴づけ、さまざまな阻害剤を比較することができる。
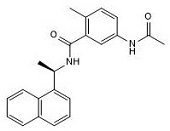
要約すると、これまでに発見された最も強力な阻害剤は、阻害剤24(化合物GRL0617としても知られている)を含むナフタレン系阻害剤である。これらの阻害剤の効力は、SARS-CoV PLproに対する阻害剤の設計に成功したことを意味し、抗ウイルス活性を備えることは、将来の他のコロナウイルスに対しても同様の特性を持つ阻害剤を設計できる可能性がある。
SARS-CoV PLpro複合体と阻害剤の研究
SARS-CoV PLproタンパク質の結晶化プロセスは難しい技術であるため、阻害剤を用いてこの酵素を結晶化することはほとんど不可能である。それでも、ロボット技術を使用することで、タンパク質-阻害剤複合体の結晶を得ることが可能であった。その結果は、結晶化条件は阻害剤ごとに異なり、このような結合は再現性がなく、新たに開発された阻害剤を使用したSARS-CoV PLproの結晶化には適用できないことがわかった。
SARS-CoV PLproとCOVID-19の関係
SARS-CoV-2 PLproの定義
SARS-CoVでは、PLpro(Papain-Like protease)タンパク質がウイルス複製の重要な役割を果たしているが、SARS-CoV-2(COVID-19パンデミックを引き起こした新型コロナウイルス)でも、ウイルス複製の機能を担い、ウイルスの拡散を可能にするウイルスタンパク質、SARS-CoV-2 PLproが見つかった。
SARS-CoV-2 PLproとSARS-CoV PLproの構造的同等性: COVID-19対策の重要なポイント
SARS-CoV-2 PLproは、SARS-CoV PLproと83%同一であることがわかっている。さらに、両方のタンパク質の活性部位の形状は非常によく類似しており、すべての重要な触媒残基は2つのプロテアーゼで実質的に変化していない。この2つのタンパク質がほとんどの構造上の特徴を共有していることは、SARS-CoV PLpro用に開発された阻害剤はSARS-CoV-2 PLproにも有効で、その結果、SARS-CoV-2のプロテアーゼ活性と細胞内での複製能力を阻害する可能性が高いことを意味する。言い換えれば、2つのウイルスプロテアーゼの構造的類似性により、SARS-CoV PLpro用にすでに開発された阻害剤を、COVID-19を引き起こすコロナウイルスに関連するタンパク質に対して使用し、同じ目的のために他の阻害剤を合成する際の参考にすることができる。
SARS-CoV-2 PLpro阻害剤
15年前から、多くの研究がPLpro阻害剤の開発に焦点を当ててきた。その結果、天然元素からナフタレン化合物まで、幅広い反応性化合物が発見された(表参照)。これらの種類の中で、ナフタレン化合物は、プロテイナーゼ阻害の有効性が最も注目されている阻害剤のグループである。
最近の研究では、GRL0617(SARS-CoV PLproの最も強力な阻害剤の1つとして前のセクションですでに説明したナフタレン阻害剤グループの化合物)がSARS-CoV-2 PLpro活性をIC50 2.2±0.3 μMで阻害できることがすでに明らかになっているが、これはSARS-CoV阻害剤24がSARS-CoV PLproに対して開発した効果(0.6±0.1 μM)よりも有意に低い。その結果、GRL0617は、CoV PLproの最も有望な阻害剤の1つとして、また、両方のタンパク質の原子レベルでの阻害機構を研究するための興味深い手段として言及されている。
脚注
参照項目
- 3C様プロテアーゼ - コロナウイルスに見られるポリタンパク質を切断するシステインプロテアーゼ